
服務熱線:1751168309217511683092
服務熱線:1751168309217511683092
養殖魚類低溫貯藏品質下降的主要原因是嗜 冷腐敗微生物的生長繁殖。 在有氧冷藏條件下,假 單胞菌多為淡水魚的特定腐敗菌。 胥亞夫等發現 水產品中主要腐敗菌為假單胞菌屬細菌, 包括熒 光 假 單 胞 菌、 腐臭假單胞菌和邊緣假單胞菌 。 Parlapani 等報道海鯛中的腐敗微生物為假單胞 菌屬,特別是熒光假單胞菌。 楊憲時等研究羅非 魚冷藏過程菌相的變化,發現在 0~10 ℃冷藏過程 中假單胞菌為優勢腐敗菌, 熒光假單胞菌為最優 勢種群。 郭全友等對養殖大黃魚的菌相研究中, 鑒定出腐敗希瓦氏菌和假單胞菌是低溫貯藏大黃 魚的優勢腐敗菌。
1 材料與方法
1.1 材料與試劑
PF01、PF06 和 PF07 為冷藏大黃魚中的優勢 腐敗菌,PF10 為大菱鲆的特定腐敗菌,NCBI 登錄 號 分 別 為 KU173826 、 KU173831 、 KU173832 、 KR706375,PFuk4 為標準菌株, 購買于微生物菌種保藏中心。 LB 肉湯、胰酶大豆肉湯(TSB)、營養瓊 脂(NA),泳動性培養基(胰蛋白胨 1%,NaCl 0.5%, 瓊脂 0.3%), 均購買于青島海博生物有限公司; SYTO9,Thermofisher,Waltham,MA,USA;96 孔 細 胞培養板,無錫 NEST 生物技術有限公司。
1.2 儀器與設備
SW-CJ-2FD 超凈工作臺,蘇凈集團蘇州安泰 空氣技術有限公司;立式壓力蒸汽滅菌器,上海博迅實業有限公司醫療設備廠;酶標儀 VICTOR X, 美 國 Perkin Elmer 公 司;移 液 器,德 國 Eppendorf 公司; 熒光顯微鏡 LEICA DM4000, 德國萊卡公 司。
1.3 方法
1.3.1 活化培養
將-80 ℃貯藏的 5 株熒光假單 胞菌甘油菌接種于 LB 培養基中 28 ℃,200 r/min 過夜活化后,于 LB 培養基中二次活化。 離心收集 菌體, 用無菌生理鹽水將菌濃度調整到 108 CFU/ mL 待用。
1.3.2 生長曲線測定
將 5 株熒光假單胞菌分離 株過夜活化菌液稀 釋 10 倍 后 按 1%接 種 于 含 有 200 μL TSB 培養基的 96 孔板中,置于 28 ℃下培 養,每隔 3 h 用酶標儀測定 600 nm 處吸光度。
1.3.3 結晶紫法定量檢測生物被膜
將過夜活化 的 細 菌 菌 液 稀 釋 10 倍 后 按 1%接 種 量 接 種 于 TSB 培養基, 并以 200 μL 分裝到 96 孔板中,28 ℃靜置培養 3,6,9,12,18,24 h 后, 采用結晶紫法 測定生物被膜形成量。 根據 Djordjevic方法略做 修改,去除孔板中菌液,每孔用 250 μL PBS 緩沖 液輕輕漂洗 3 次,干燥后,加入 200 μL 0.2%的結 晶 紫 溶 液,染 色 15 min 后 清 洗,干 燥,最 后 加 入 200 μL 33%冰乙酸, 用酶標儀測量其 590 nm 處 吸光值。
1.3.4 胞 外 多 糖 (EPS) 含 量 測 定
參 考 Harimawan 等的方法略作修改提取胞外多糖。 具體 為培養 12,18,24 h 后的 10 mL 菌液,4 ℃離心(5 000×G,20 min), 去 上 清 , 然 后 用 磷 酸 緩 沖 液 (Phosphate Buffered Saline,PBS)重懸菌體,在 80 ℃水浴 30 min,4 ℃離心 (15 000×G,30 min),用 0.22 μm 濾膜過濾,得到上清液用苯酚硫酸法測定 胞外多糖含量。
1.3.5 珠渦流法
在 1.3.1 節所述的細菌菌懸液中加入不銹鋼片 (10 mm ×10 mm) 和蓋玻片(22 mm × 22 mm)。在 28 ℃靜置培養 1 h。參考 Nguyen 等珠渦流法對粘附在不銹鋼片表面的細菌進行 計數。具體方法如下,用 PBS 緩沖液輕輕洗去不銹 鋼片表面的浮游菌, 然后放于裝有 10 mL 無菌生 理鹽水的離心管中,加入適量玻璃珠,劇烈渦旋 3 min 后,梯度稀釋計數。
1.3.6 熒光顯微鏡觀察
熒光顯微鏡觀察參照 Bagge 等的方法。 具體如下:用滅菌 PBS 洗去蓋 玻片上浮游菌, 放入 2.5%的戊二醛中固定 1.5 h, 用 PBS 洗去戊二醛,烘箱干燥后,滴加 70 μL 0.1 μL/mL SYTO9 于蓋玻片上, 均勻覆蓋于表面,避 光孵育 20 min,吸去多余的染料后,放置熒光顯微 鏡下(40×)觀察。
1.3.7 泳動性測定
參考 Sperandio 等的方法, 泳動瓊脂培養基凝固后,將 5 μL 過夜培養的菌液 滴在平板中心,吸干后,移至 28 ℃培養箱中靜置 培養 24 h。
1.3.8 蛋白酶活性測定
參照 SB/T 10317-1999 標準中福林酚法測定蛋白酶活力方法,在 40 ℃ 下每分鐘水解酪蛋白產生 1 μg 定義為 1 個蛋白 酶活力單位。
1.3.9 嗜鐵素檢測
參考文獻,制作 CASAD 嗜鐵素檢測平板,利用 CASAD 嗜鐵素平板法可檢 測總的嗜鐵素, 將過夜培養的菌液離心取上清 5 μL 加入平板中,置于 28 ℃培養 24 h 后觀察。
1.3.10 生 物 報 告 菌 檢 測
信 號 分 子 參 考 Ravn 等方法并略作修改。 報告菌紫桿菌 CV026 活化 后,取 5 mL 加入到 50 mL LB 瓊脂中,混合均勻后 倒平板,待冷卻后,平板中心打 孔,加 入 菌 液,28 ℃培養,觀察顏色變化。
1.3.11 統計學處理
每組樣品設 3 個重復,結果 取其平均值。采用 Microsoft Excel 和 Origin 8.5 進 行數據處理和 作圖, 并利用 SPSS21.0 的 ANOVA 進 行 方 差 分 析 P<0.05 表示有統計學意 義。
2 結果與分析
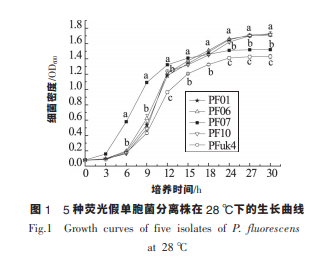
研究測定了 5 株熒光假單胞菌分離株在 28 ℃下靜置培養的生長曲線。 如圖 1 所示,熒光假單胞菌在 3 h 到 12 h 期間生長迅速,尤其是 PF07 在 對數生長期生長最快,PF01、PF06 和 PF10 次之, 且這 3 株菌的生長基本保持一致,PFuk4 生長最慢。 在 18 h 后 PF07 和 PFuk4 生長緩慢,在培養 24 h 進 入穩定期,而其余 3 株菌依然在逐漸增長,在 27 h 進入穩定期。 PF01、PF06、PF07、PF10 和PFuk4 5 株菌在穩定期的細菌量分別達到 1.7,1.7,1.5,1.7 和 1.4(OD600),其中 PF01、PF06 和 PF10 的細菌量 最高,PFuk4 的菌量最少。
熒光假單胞菌在 96 孔塑料孔板上也能形成 被膜, 其中培養 3 h 時 5 株菌的生物被膜量基本 保持一致(P>0.05)。 隨著細菌生長,熒光假單胞菌 的生物被膜量也開始顯著增加 , 培 養 至 12 h, PF01、PF06、PF07 和 PF10 的 生 物 被 膜 量 達 到 最 高,分別為 1.71,1.72,2.24 和 1.61(OD600),而 PFuk4 在 18 h 達到最高,為 2.50,隨即又開始下降。 在熒 光假單胞菌分離株中 PF07 和 PFuk4 的生物被膜形 成能力較強,且 PF07 被膜形成周期較短。 研究表 明,熒光假單胞菌具有較強的生物被膜能力,而且 不同來源的熒光假單胞菌生物被膜形成能力差別 較大。 Sim觛es 等報道熒光假單胞菌模式菌株形 成生物被膜的能力高于從乳制品加工環境中分離 得到的分離株。 課題組也發現不同來源的銅綠 假單胞菌形成生物被膜的能力也有所不同。 在食 品接觸表面的腐敗致病菌的粘附及生物被膜形 成,增加產品加工過程的交叉污染,縮短貨架期, 給食品產業帶來嚴峻挑戰。
3 結論
研究表明,5 株熒光假單胞菌都能在試管和 塑料孔板中形成大量生物被膜, 且能較快地粘附 于不銹鋼片與玻璃片表面。 熒光假單胞菌具有較 強的泳動性、蛋白酶活性及產嗜鐵素,其中 PF07 較強的生物被膜形成能力和致腐表型, 可能與高 活性的 AHLs 存在內在關聯。

